В рамках работы по русификации обзорного каталога исследований по мышечной дистрофии, проводимых по всему миру, на сайте МойМио в разделе Каталог исследований по МДД опубликована статья посвященная исследованию сосудистой доставки к конечностям гена микродистрофина без введения большого объема жидкости или большого давления для лечения миодистрофии Дюшена.
Успешная генотерапия миодистрофии Дюшена потребует трансдукции мышцы, используя ген замены, по-видимому мини — или микродистрофин или суррогатный ген. Успех у mdx мыши с поставкой через вены хвоста известен, но остается еще много вопросов до перехода к клиническим испытаниям. В этом исследовании мы сосредоточились на сосудистом способе доставки, которая может быть применена в клинических испытаниях, для достижения клинически значащего результата, предоставляя безопасную доставку вируса, и используя невысокую дозу, что снимает производственные ограничения для rAAV векторов. Мы рассуждали, что региональная сосудистая доставка к нижним конечностям через бедренную артерию могла потенциально оправдать эти надежды.
Наше исследование устанавливало строгие критерии для трансгенеза с определением возможностей для клинического применения, используя подход и серотип AAV, который потенциально ускорит трансдукцию мышц, и как следствие приведет к улучшению качества жизни у детей с миодистрофией Дюшена, с возможностью применения для лечения других мышечных дистрофий.
Сосудистая доставка к конечностям гена микродистрофина без введения большого объема жидкости или давления для лечения миодистрофии Дюшена.
Louise R Rodino-Klapac1,2, Paul ML Janssen3, Chrystal L Montgomery1,2, Brian D Coley4, Louis G Chicoine1,2, K Reed Clark1,2 and Jerry R Mendell1,2*
- * Corresponding author: Jerry R Mendell MendellJ@ccri.net
Author Affiliations
1 Center for Gene Therapy, Columbus Children’s Research Institute, Columbus Children’s Hospital, 700 Children’s Dr., Columbus, Ohio, 43205, USA
2 Department of Pediatrics and Neurology, The Ohio State University, Columbus, OH 43210, USA
3 Department of Physiology and Cell Biology, 304 Hamilton Hall, College of Medicine, The Ohio State University, Columbus, OH 43210, USA
4 Department of Radiology, Columbus Children’s Hospital, 700 Children’s Dr, Columbus, OH 43205, USA
Journal of Translational Medicine 2007, 5:45 doi:10.1186/1479-5876-5-45
Оригинал статьи: http://www.translational-medicine.com/content/5/1/45
Миодистрофия Дюшена- наследственное заболевание с рецессивным Х сцепленным наследованием, вызванное мутацией в гене дистрофина, а следовательно перспективное для разработки генной терапии. Мы закончили изучение обнаружения подходов генной терапии для проведения клинических исследований, руководствуясь следующими критериями:
1. Доставка через региональные сосуды во избежание диссеминации вируса по всему организму.
2.Подход потенциально облегчающий поступление вируса для наиболее эффективного преобразования скелетных мышц.
3. Использование доз вируса в соответствии с существующими ограничениями.
4. Но в то же время достичь клинически выраженного результата.
Предпосылки
Миодистрофия Дюшена одно из наиболее часто встречающихся наследственных прогрессирующих мышечных заболеваний, выявляемых в детском возрасте и поражающих 1 из 3500 мальчиков. Ограниченные возможности лечения, представленные в основном использованием кортикостероидов, продлевают способность к передвижению не у всех пациентов. Стратегии комплексного лечения находятся в процессе разработки. Они включают такие методы, как пропуск экзона, преждевременную остановку кодонов и фармакологические препараты, направленные на развитие мышц. Обнадеживающие данные получены при замещении гена дистрофина в эксперементальных исследованиях у мышей и собак. Выводы , извлеченные из данных исследований, в настоящее время говорят об ограничении использования AAV для доставки гена, введенного местно в мышцу путем инъекции. Эти исследования служат фундаментом для будущих открытий, но в настоящее время уделяется большее внимание системной доставке вируса, как методу с наиболее ожидаемым положительным эффектом, значимым для улучшения качества жизни пациентов. Доставка вируса путем инъекций в региональные сосуды определяет направление для будущих клинических испытаний.
У мышей получавших 6 или 8 серотип AAV системно было обнаружено широкое распространение микродистрофина в скелетных мыщцах. Эти предшествующие исследования интересны для проведения клинических исследований, но требуют значительной доработки. Очень большая вирусная нагрузка, необходимая для системного распространения AAV6 используемая у mdx мышей из-за сложностей с производством может существенно ограничить клиническое использование данного метода. Исследования AAV8 проводились только на новорожденных мышах, а следовательно полученные результаты могут существенно отличаться у взрослых мышей и тем более у пациентов с миодистрофией Дюшена. В дополнение ко всему сказанному, обсуждается использование AAV1 для трансформации мышечной ткани, так как получены хорошие результаты при его введении через бедренную артерии у mdx мышей. Однако Верный способ применения для проведения клинических испытаний AAV1 еще не разработан, так как испытания проводимые у мышей предполагали очень большой объем жидкости вводимый через бедренную артерию, что вызывало разрыв капилляров эндотелия и отек нижней конечности.
Методы
Животные.
Все процедуры были одобрены комитетом по использованию животных. Трех, четырех недельные mdx мыши и C57/BL10 использованы для данного исследования. 1-2х летние макаки получали ELISA для связывания антител к AAV8.
Изучение перфузии конечностей у мышей
Мыши получили кетамин и гилазон. Сбрита шерсть на левом бедре, данная область обработана 95% этанолом и раствором йода. Через небольшой разрез визуализирована бедренная связка проксимальней середины бедра, выделены бедренные сосуды. Под сосуды подведена шелковая лигатура. В сосуды установлены полипропиленовые катетеры для контроля кровотока. В артериальный катетер введено 100мкл физиологического раствора. Затем кровоток в конечности был прекращен с помощью лигатуры. Венозный катетер был закрыт.В артериальный катетер введено 1 *10^11 вирусных частиц (rAAV1, rAAV6, r AAV8) в 100 мкл физиологического раствора. Забуфференный физиологический раствор вводился со скоростью 2 мкл/ сек. Через 10 минут был открыт венозный катетер, в артериальный катетер введено 100мкл физиологического раствора. После чего катетеры удалены, кровотечение остановлено простым сжатием сосудов. Рана была ушита.
Для внутримышечных инъекций мышам проведена анастезия 1-4% изофлураном. В обе нижние конечности введено 3* 10 ^10 rAAV1, 6, 8 вирусных частиц или 30 мкл физиологического раствора с использованием инсулинового шприца.
Изучение перфузии конечностей у макак.
Макаки Cynamologous получили внутримышечно телазол (3-6 мг/кг) и интубированны. Общий наркоз в течение процедуры проводился 1-4% изофлураном. Левый крестцовый свод обрит до середины бедра , данная область обработана 95% этанолом и раствором йода. Было проведено выделение бедренной артерии и вены. Артерия была катетеризирована, через катетер введено 2 мл физиологического раствора. Произведена флеботомия, кровоток в конечности прекращен путем наложения лигатуры. Введена AAV8 или CMV или eGFP в дозе 10^13 в 2 мл буферного раствора. Циркуляция вируса происходила в течение 10минут. Гемостаз и ушивание раны как в предыдущем эксперименте.
Сборка гена микродистрофина.
Мышиный микродистрофин содержал области R4–R23/?71–78. Ядерная ДНК оптимизирована кодонами грызунов и синтезирована GenScript Inc. Она включала согласованную Kozak последовательность, SV 40 интрон и синтетический полиА сайт. Усеченный промотер/ энхансер мышечной креатинкиназы(МСК) использован для управления специфической мышечной экспрессией генов. МСК микродистрофин был клонирован AAV2 ITRs, используя Xba I рестрикционный энзим, в плазмиды pCMV?. Msc I/Sma I рестрикционный энзим был использован для подтверждения интеграции.
Продукция rAAV.
rAAV вектор синтезирован при помощи модифицированного перекрестно-упаковочного метода, в следствие чего геном AAV2 мог быть упакован в сложный AAV капсид. Продукция была проведена стандартной 3 плазмидой DNA/CaPO4 преципитационным методом используя HEK293 клетки. 293 клетки поместили в DMEM с 10% фетальной бычьей сывороткой, пенициллином и стрептомицином. Получены плазмиды: pAAV.MCK. microdys, (ii) rep2-capXмодифицированная AAV хелпер плазмиды, кодируемые серотипами 1,6 или 8 изолятов; и (iii) аденовирусная хелперная плазмида 5 типа. Проведенный флюоресцентный анализ на МСК промотер показал: МСК, 5-CCCGAGATGCCTGGTTATAATT-3; MCK 5-GCTCAGGCAGCAGGTGTTG-3; и MCK, 5-FAM-CCAGACATGTGGCTGCTCCCCC-TAMRA-3.
Анализ экспрессии генов.
Передняя большеберцовая и икроножная мышцы были взяты для исследования через 4,8, 12 недель после лечения. Мышцы помещены в трагакантовую камедь и заморожены в изопентане охлажденном жидким азотом. Криостатный срез для окраски был обработан N-terminal Manex1 первичными антителами (Developmental Studies Hybridoma Bank), первичными антителами к альфасаркогликану и бетасаркогликану в разведении 1:50 в течении 1 часа при комнатной температуре. Срезы промыты PBS трижды, каждый 20минут и заморожены. Визуализация осуществлялась после обработки Alexa 488 или Alexa 594 конъюгировании изотип- специфическими противомышинными антителами в разведении 1:300. Флюоресценция микродистрофина наблюдалась при освещении светом с длиной волны 488нм для ФИТЦ с использованием Zeiss Axioskop2 Plus микроскопа , изображения были захвачены с помощью Zeiss AxioCam MRC5 camera.
Блоттинг
Срезы тканей C57/BL10, mdx мышей из контрольной и группы получавшей лечение гомогенизированы. После гомогенизации образцы центрифугированы в течении 5 минут. Проведен электрофарез с 3-8% полиакриловым гелем (NuPAGE, Invitrogen) 16 часов и затем на PVDF мембране 1 час при 4°С Мембрана обработана обезжиренным5% сухим молоком в TBST (100 mM Tris-HCL, pH 8.0, 167 mM NaCL, 0.1% Tween) 1 час и инкубирована с первичными моноклональными антителами Manex1a (Developmental Studies Hybridoma Bank) в разведении 1:200 и миоспецифичными моноклональными антителами (Vector Labs).
Морфометрия.
Централизованный счет ядер выполнен в срезах передней большеберцовой мышцы при окраске гемотоксилином, эозином у 3-х месячных mdx животных получавших лечение. Другая конечность служила для контроля.На 5 случайных полях 12 срезов каждой мышцы было определено количество ядер.Также был определен диаметр мышечных волокон и проведена окраска сукцинилдегидрогеназой для определения митохондриальной активности в волокнах. Контрольную группу составляли мышцы другой конечности.
Мышечная сила и прочность при эксцентричном напряжении.
У 8 недельных мышей взят длинный разгибатель пальцев и помещен в насыщенный кислородом раствор Krebs-Henseleit (95% O2/5% CO2 (pH 7.4) 118 mM NaCl, 25 mM NaHCO3, 5 mM KCl, 1 mM KH2PO4, 2.5 mM CaCl2, 1 mM MgCl2, 5 mM Glucose) при 30°С. Один конец мышцы был прикреплен к датчику силы, а другой к высокоскоростному линейному сервоуправляемому мотору. Мышечная длина изменялась в заданных значениях. Электрическая стимуляция проводилась через два платино-иридиевых электрода с каждой стороны мышцы. Мыщцы подвергнуты трем изометрическим сокращениям каждое по 500мс с минутным перерывом между стимуляцией.. после 5 минутного перерыва мышцы были подвергнуты 4 сериям изометрического напряжения с возрастающей частотой 50, 80, 120, 150Гц каждое по 500мс с минутным перерывом между стимуляцией. После 10 минутного перерыва мышцы были подвергнуты эксцентричному сокращению из серии 10 изометрических 700мс напряжений с 2-х минутными интервалами, с 10% удлинением мышц при достижении максимальной силы. После окончания сокращений мышцы приведены к первоначальной длине. Для сравнения силы, она оценивалась в отношении к площади поперечного сечения волокна.
Статистический анализ.
Данные анализированы стандартными статистическими методами. Для определения статистической достоверности был использован t-тест и десперсионный анализ. Уровень достоверности p = 0.05.
Результаты
Конструкция rAAV для переноса генов микродистрофина.
Первой целью была идентификация оптимального rAAV серотипа, обеспечивающего эффективный сосудистый выход в подлежащую ткань мышцы; вирусных доз, и использования объемов и давления перфузии, адекватных клиническому применению. Исходя из данных предыдущих работ, серотипы AAV 1, 6 и 8, рассматривались в качестве ведущих кандидатов. Генетический материал состоял из гена микродистрофина под контролем обрезанного промотора/энхансера MCK (563 bp), чтобы достигнуть в мышцах определенной экспрессии гена, и избежать разнородной трансгенной экспрессии в нежелательных типах клеток, помогая уменьшить иммунный ответ. Конструкция микродистрофина, (R4–R23/?71–78), идентичная описанному Харпером и др. [20] и позже используемая Йошимура и др. [12], содержит N-окончание, связанную актином область, четыре повторения спектрина и богатую цистеином область (Рис. 1А). Мышиная КОМПЛЕМЕНТАРНАЯ ДНК синтезировалась с использованием оптимизации кодонов. Несколько модификаций добавлены для достижения максимальной экспрессии гена: (1) интрон из SV40 (120 bp), для стабильности и увеличения ядерного экспорта [23]; (2) маленький синтетическое полиаденилирование для минимизации размера конструкции и (3) включение согласованной последовательность Kozak (CCACC) как сигнал инициирования трансляции. Все векторы были подготовлены, используя протокол трансфекции ДНК плазмиды без аденовирусов и очищены Векторные титры генома измерены Taqman Q-PCR как ранее описано нашей группой. [22]
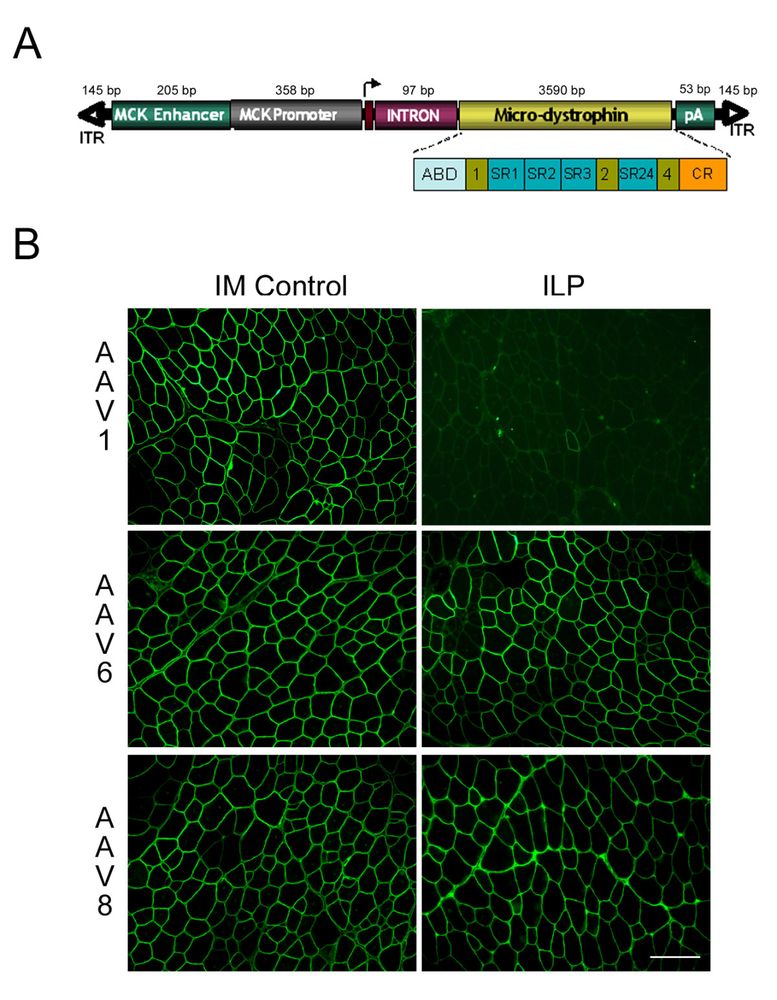
Рисунок1. (A) Схематичный микродистрофин. Обрезанный промотор MCK (563 bp) используется, чтобы контролировать экспрессию гена. Также маркированный интрон SV40 (97 bp) и синтетическое полиаденилирование (53 bp). 3 590 bp мышиного микродистрофина изображается подробно. ABD- полный актин-обязательная область, стержни 1, 2 и 4 (зеленые области)- фланговые области спектрина (СЭР синие области). Богатый цистеином дистрогликан обязательная область обозначается оранжевым. AAV2 ITR показанны стрелками. (B) Иммунофлюоресценция микродистрофина у mdx мыши в передней большеберцовой мышце . rAAV1, 6, или 8. (1011 vg) был введен внутримышечной инъекцией (Контроль IM) или ILP через бедренную артерию 3-4-недельных mdx мышей. Представленны срезы передних большеберцовых мышц (12 гм) пок 4-недельных животных получавших лечение (срезы 8 и 12 недельных животных выглядели подобно). Срезы были окрашены N-терминальными антителами к дистрофину Manex1a.
Эффективные результаты доставка rAAV6 и rAAV8 к конечности мыши
rAAV1, 6, или 8 вектор, ген микродистрофина, MCK введен в дозе 10^11 вирусных частиц в стерильном буферном солевом растворе в бедренную артерию левой задней конечности 3-4-недельных mdx мышей. Детали размещения катетера описаны в методах, но определенные шаги могут быть акцентированы. Анатомические ограничения у мыши привели к размещению катетера ниже слияния глубокой бедренной артерии, основного источника кровоснабжения бедра и мышц. До введения вектора,промыт артериальный катетер 100мкл стерильного физиологического раствора. Впоследствии шелковые лигатуры были сжаты, чтобы предотвратить весь кровоток в конечности (изолированная перфузия-ILP конечности) ближайший к месту введения катетера. Немедленно после остановки кровотока, вектор медленно вводится (более чем 60 — 80 секунд), чтобы минимизировать капиллярный разрыв. Экспрессия гена микродистрофина была определена количественно иммунным окрашиванием поперечных разрезов мышцы, используя моноклональные антитела (Manex-1a). Выраженная экспрессия микродистрофина наблюдалась в передней большеберцовой мышце и длинном разгибателе пальцев мышей, получающих rAAV6 и rAAV8. Количество волокн содержащих микродистрофин у мышей получавщих rAAV8 94.5 ± 0.9 % (1 месяц), 91.3 ± 3.1 % (2 месяца), и 89.6 ± 1.6 % (3 месяца); и rAAV6- 87.7 ± 6.8 % (1 месяц), 78.9 ± 7.4 % (2 месяца), и 81.2 ± 6.2 % (3 месяца) (Рис. 1B). В икроножной мышце определены более низкие уровни экспрессии во всех 3 моментах времени 6.7 ± 0.7 % для rAAV8, и 17.6 ± 1.4 % для rAAV6. Это, вероятно, вызвано размещению катетера и анатомии сосудистой сети задней конечности мыши. У мыши бедренная артерия раздваивается на уровне колена с одним отходящей артерией, снабжающей кровью переднюю большеберцовую мышцу и длинный разгибатель пальцев и две меньших артерии, которые снабжают кровью икроножную и комбаловидную мышцы. Бедренный катетер был установлен выше колена и поставка вектора была направлена к более доступной артерии, к передней большеберцовой мышце. В отличие от rAAV8 и rAAV6, сосудистая поставка rAAV1 не эффективна в мышцах нижней конечности 0.9 ± 0.3 % (1 месяц), 2.1 ± 0.8 % (2 месяца), и 2.1 ± 0.7 % (3 месяца) (Рис). Различия в дозе, объеме инъекции, и давлении лучше всего объясняют различные результаты (см. обсуждение).
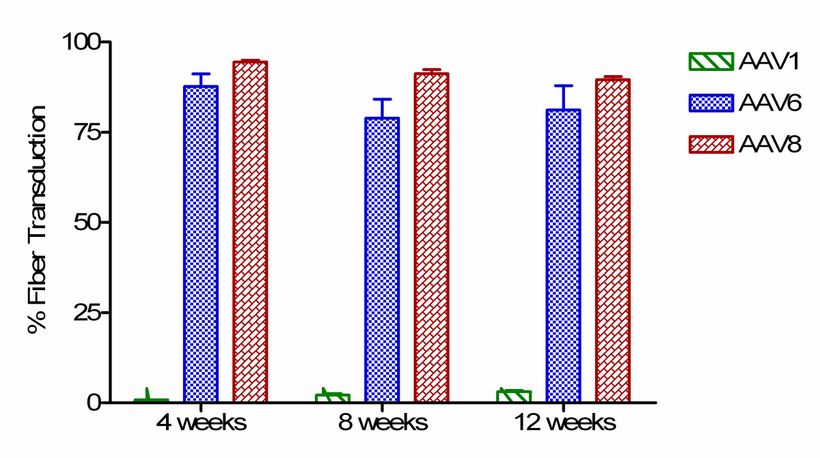
Рисунок2. Определение количества экспрессии микродистрофина. Распределение микродистрофина в срезах мышцы. Уровень экспрессии микродистрофина был определен количественно- визуальным количеством волокон из 4 случайных областей (число положительных волокон/общее количество волокон (n = 4–8 за группу).
Связь уровня экспрессии микродистрофина с выраженностью патологии
Главная волна некроза и регенерации у mdx мыши встречается приблизительно в 3 — 4 недельном возрасте, совпадающего с нашим лечением. Максимальная экспрессия гена rAAV векторов встречается приблизительно в 4х недельном возрасте. Таким образом положительные эффекты достаточно скромные. Количественная оценка ядер была определена в передних большеберцовых мышцах мышей получавших rAAV6 или rAAV8 и соотносится с контрольными группами. Ядра были меньше на 25 % (rAAV8 – 65.3 ± 3.3 %, rAAV6 – 64.05 ± 3.4 %, у mdx – 86.16 ± 0.9 %; P <0.0001; Рис3Е.),как тражение рецидивирующих циклов регенерации и дегенерации только между временем экспрессии гена и вскрытием трупа (2 месяца объявляют о переносе генов). Подробный морфометрический анализ диаметра волокна показал, что у rAAV6 и у групп получавших rAAV8 обнаружено увеличение диаметра мышечного волокна по сравнению с контрольными группами. Поперечные срезы групп получавших лечение и группы контроля с медленными волокнами (тип 1), волокнами с быстрым окислительным типом гликолиза (тип 2A) и быстрым гликолизом (тип 2B) , с медленными волокнами, показали самое большое увеличение (Рис. 3E ). Существенные различия в диаметре волокна (3F) были замечены у животных, получавших rAAV8. по сравнению с другими группами [тип 1-39.2 ± 0.4 (rAAV8), 30.2 ?m (mdx); тип 2A — 48.2 ± 0.7, 38.9 ± 0.5 ?m; тип 2B — 52.4± 0.8 против 47.1 ± 0.6 ?m, где P <0.0001], в то время как у животных, получавших AAV6, показано существенные различия для типа 1 (33.2 ± 0.3 ?m; P <0.0001) и типа 2A (40.8 ± 0.5 ?m; P <0.01) волокон. Самые большие различия и для rAAV8 и для rAAV6 были продемонстрированы в типе 1 волокон с 23%-ым и 10%-ым увеличением диаметра волокна, соответственно. Эти различия далее демонстрируются в иллюстрации, где показано распределение частоты диаметров волокна, стратифицированных типов волокон, увеличение диаметра и для rAAV8 и для rAAV6. Дисперсионный анализ был выполнен для всех наборов данных, значения для обеих групп по сравнению с контрольными группами mdx P <0.0001.
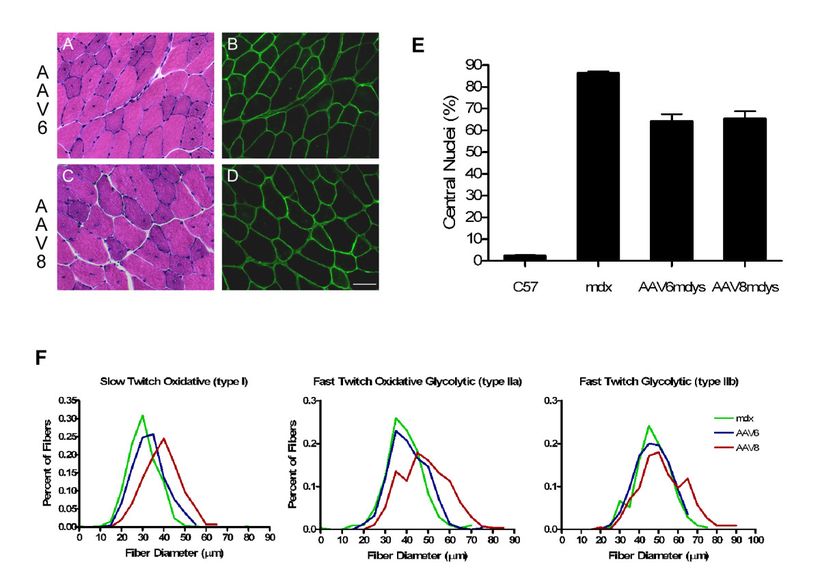
Рисунок 3. Уровни экспрессии микродистрофина пропорционален снижению патологии. ( Гематоксилин, эозин) Анализ показал сокращение централизованных ядер, признаков патологии при миодистрофии. (A, C) Окрашивание срезов мышц животных получавших rAAV6 и rAAV8 (8 недель).(B,D)обнаружение иммунофлюоресценции микродистрофина в последовательных срезах мышц животных получавших rAAV6 и rAAV8 соответственно. (E) Экспрессия микродистрофина заканчивается 25%-ым уменьшением в централизованных ядрах. (F) распределение результатов экспрессии микродистрофина, значительное увеличение диаметра волокон типа 1, IIa, и IIb для mdx мышей получавших rAAV6 и rAAV8 по сравнению с контрольными группами mdx. (n = 5-6 за группу) (p <0.0001 анализа АНОВОЙ).
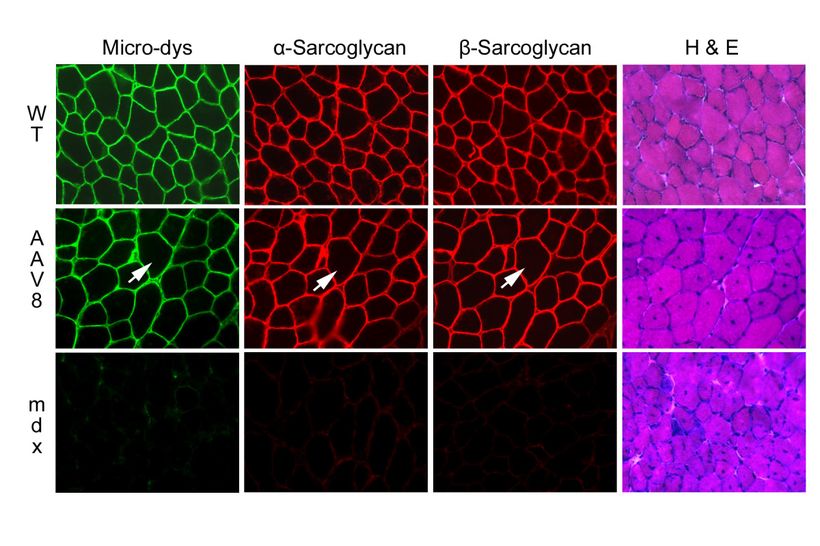
Рисунок4. Компоненты дистрофин-связанного белкового комплекса восстанавливаются у mdx мышей, получавших rAAV. Мыши mdx, получавшие rAAV8 (8-недель) были окрашены manex1a антителами дистрофину, ?-саркогликану, ?-саркогликану, и гематоксилином, эозином. Волокна мышцы показали восстановление экспрессии ?-саркогликана и ?-саркогликана (отсутствие окрашивания у mdx мышей не получавших лечения — нижний ряд).
Связь экспрессии микродистрофина и увеличения силы mdx животных
Чтобы проверить, привела ли экспрессия микродистрофина к улучшению на функциональном уровне, сила мышц mdx мышей получавших rAAV бала оценена по сравнению с mdx мышами не получавшими лечение и контрольными группами дикого типа C57/B10. Исследователи анализировали in vitro измерений силы длинного разгибателя пальцев. rAAV8-и rAAV6-леченные мышцы показали 50%-ое увеличение максимальной силы изометрического сокращения по отношению к площади поперечного сечения мышцы (rAAV8 146.5 ± 7.6 mN/mm2, rAAV6 142.2 ± 14.8 mN/mm2 контрольная mdx группа 109.5 ± 5.1 mN/mm2; P <0.05) (Рис. 5A). Увеличение развитой силы не достигало уровня контрольных групп дикого типа C57BL/10 (198.3 ± 11.9 mN/mm2; P <0.05) (Рис. 5A). После исследования силы мышцы были подвергнуты механическому повреждению повторными эксцентричными сокращениями. Экспрессия микродистрофина у мышей получавших rAAV8 и rAAV6 определяла защиту от повреждения при эксцентричном сокращении. Леченные мышцы были значительно более устойчивы к вызванному сокращением повреждению по сравнению с нелеченным (p <0.05 (Рис. 5B) Эти данные демонстрируют, что rAAV6 или rAAV8 введенные через сосудистую сеть, приводит к значительному увеличению силы изометрического сокращения и значительно уменьшает повреждение при эксцентричном сокращении.
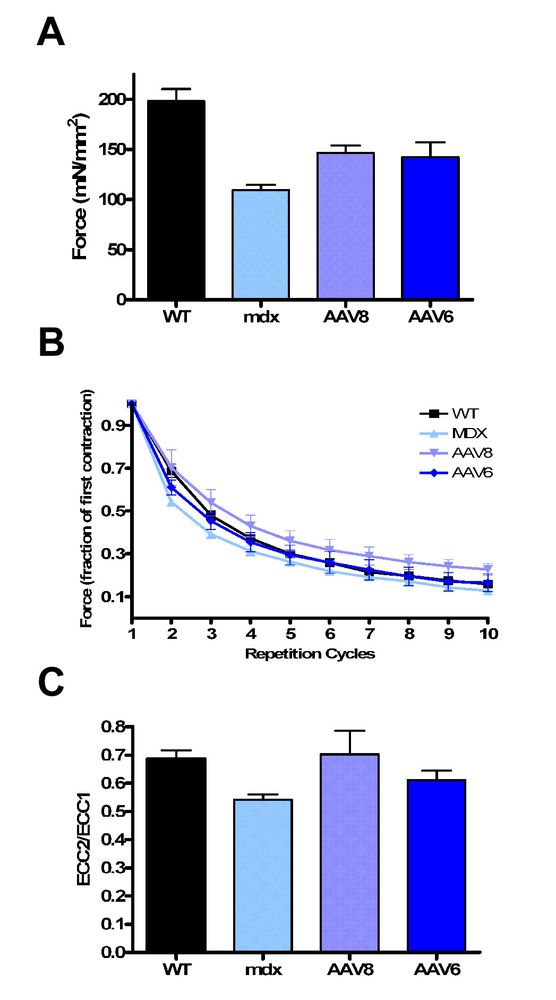
Рисунок 5.Перенос генов микродистрофина вызывает улучшения силы значительно. (A) Длинный разгибатель пальцев продемонстрировал значительное максимальное увеличение силы (mN/mm2) по mdx-нелеченным мышцам (p <0.05), но не достигал уровней дикого типа C57BL/10 (ВЕС) контрольной группы (P <0.05 (B) мышцы были подвергнуты 10 циклам изометрического возбуждения с частотой 150 гц с удлинением 10 %, чтобы вызвать повреждение в течение 100 миллисекунд. rAAV8 и rAAV6 показанную значительную защиту мышц EDL от повреждения по сравнению с mdx-нелеченными группами во время первых двух циклов (p <0.05), после чего все 4 группы показали наличие повреждений, что различия больше не могли быть определены. Не было никакой значительной разницы между rAAV8 или rAAV6 (C)
Сосудистая доставка rAAV8 у маккак cynamologous заканчивается широко распространенной экспрессией гена в нижней конечности
Цель состоит в том, чтобы применить преклинические результаты исследования к клинической практике. Не принадлежащие к человеческому роду приматы (NHP) представляют собой модель с анатомическими параметрами, близкими к людям, и позволяют установить приблизительные диапазоны дозирования, подобные маленькому ребенку. Доза, примененная к макакам, была пропорциональна (основанный на весе животных) дозе у мышей (5 ? 1012 vg/kg у мыши и 2 ? 1012 vg/kg в NHP).В левую заднюю конечность двух cynamologous макак (4-5-килограммовые животные) было введено приблизительно 1013 вирусных частиц (2 ? 1012/кг) rAAV8. CMV, eGFP в 2 мл физиологического раствора, используя изготовленный на заказ, рентгенконтрастный катетер, который был установлен ближе к артериальной бифуркации на уровне колена. Процедура схожа с исследованием у мышей хотя, бедренная вена не была катетеризована для этого эксперимента.Через три недели проведено исследование мышц нижней конечности включая TA, EDL, gastrocnemius и soleus, чтобы сравнить с исследованиями mdx мышей. GFP визуализировался прямой флюоресценцией, и продемонстрировал широко распространенную экспрессию гена во всех мышцах [средний gastrocnemius — 63.8 ± 4.9 %, боковой gastrocnemius — 66.0 ± 4.5 %, EDL — 80.2 ± 3.1 %, soleus — 86.4 ± 1.9 %, TA — 72.2 ± 4.0. Данные были получены, при оценке 4 областей из 4-6 блоков мышцы для каждой мышцы, охватывающей ближайшие, центральные, и дистальные области, в общей сложности 16-24 — 20? области (область = 0.349 mm2). Эти данные демонстрируют, что rAAV8 эффективен при получении доступа к основной мускулатуре у примата и, кажется, идеальный серотип для поставки к мышцам через сосудистую сеть, используя низкий объем, низкое давление, и низкую дозу.
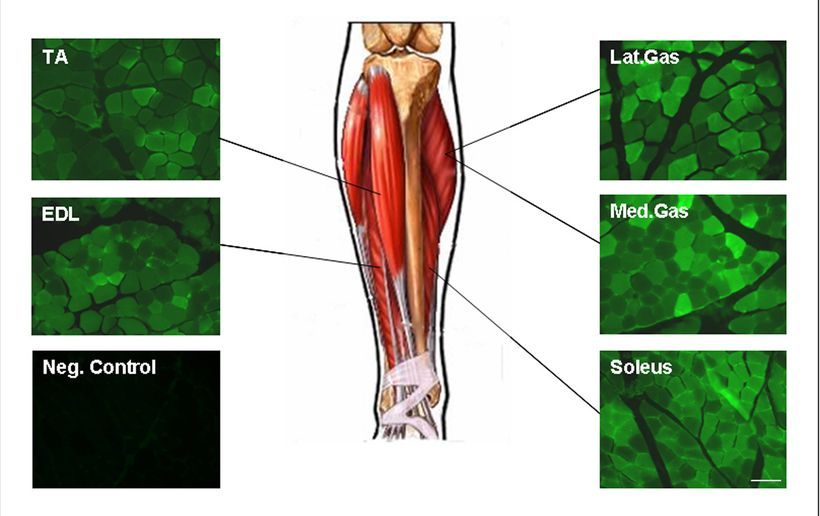
Рисунок 6. Изолированная перфузия конечности при введении rAAV8. CMV.eGFP у cynomologous макаках (4-5 кг) заканчивалсь широко распространенной экспрессией гена в мышцах нижней конечности. 2 ? 1012 vg/kg (объем на 2 мл) было введено через бедренную артерию. Во время 10-минутного прекращения кровотока происходило связывание вектора. Оценка результатов проводилась через 3 недели.
Обсуждение
Успешная генотерапия миодистрофии Дюшена потребует трансдукции мышцы, используя ген замены, по-видимому мини — или микродистрофин или суррогатный ген. Успех у mdx мыши с поставкой через вены хвоста известен, но остается еще много вопросов до перехода к клиническим испытаниям. В этом исследовании мы сосредоточились на сосудистом способе доставки, которая может быть применена в клинических испытаниях, для достижения клинически значащего результата, предоставляя безопасную доставку вируса, и используя невысокую дозу, что снимает производственные ограничения для rAAV векторов. Мы рассуждали, что региональная сосудистая доставка к нижним конечностям через бедренную артерию могла потенциально оправдать эти надежды.
Первый шаг в этом процессе — идентифицировать адекватный серотип AAV, который эффективно пересечет сосудистый барьер,несмотря на низкое давление и низкий объем. Есть сторонники использования AAV6 и AAV8; однако, вирусная нагрузка, сосудистые факторы, и возраст животных устраняют прямые сравнения между этими серотипами в изданной литературе. AAV1 также продемонстрировал активность после сосудистой поставки у мышей, но условия для поставки не могут быть сопоставленны с AAV6 и AAV8. Для клинических испытаний мы ожидаем использование региональных сосудистых доступов для поставки вируса. Бедренная артерия, снабжающая главные группы мышцы в нижней конечности, представляет собой основной сосудистый доступ для введения вируса, и первой целью стало сравнение серотипов AAV (AAV1, AAV6, и AAV8),с тем же самым трансген (модифицированный микродистрофин), под контролем того же самого промотора (MCK), используя идентичные условия поставки, адекватные клинические условия. Результаты были однозначны. AAV6 и AAV8 показали примерно одинаковые результаты для региональной сосудистой поставки к нижней конечности, которые поразительно отличалась от AAV1. Мы нашли широко распространенную экспрессию микродистрофина в TA и EDL через 1, 2, и 3 месяца, указывающую на перенос генов с незначащими различиями в количественной оценке преобразованных мышечных волокон (rAAV8> 90 % и rAAV6> 80 %). Эти полезные действия трансдукции через сосудистый путь поставки продемонстрировали значительное увеличение силы и защиты от вызванного сокращением повреждения.
Эффективность трансдукции для rAAV1 была резко снижена (<3 %). Мы приписываем различия между результатами AAV1 и таковыми из предшествующих исследований объему перфузата [1 мл против 100 мкл (текущее исследование) на 20-граммовую массу тела] и скорости введения [ 10-39 секунд против 60-80 секунд (текущее исследование)]. Используя AAV1 для сосудистого переноса генов, Gonin и др. продемонстрировал капиллярный разрыв в пределах эндомизия, что скорее всего повлияло на высокий показатель экспрессии гена. Они также наблюдали отек оконечности. Хотя мы непосредственно не исследовали капиллярное русло в наших исследованиях, мы предполагаем, что сосудистая целостность была сохранена: 1) не было отеков конечностей у наших мышей; 2) AAV1, AAV6, и AAV8 продемонстрировали равные полезные действия трансдукции после внутримышечных инъекций- 94%-ая экспрессия гена, внушительные различия были замечены после сосудистого переноса генов, или 3) подавляющее закрепление в пределах сосудистой сети rAAV1.
В ходе тестирования серотипов для поставки микродистрофина, мы поставили цель выявить подход переноса генов, который мог потенциально быть приспособлен к клинике. У мыши мы использовали изолированную перфузию конечности, предварительный прилив крови и солевого раствора до трансгенеза, время с остановленным кровообращением и, прилив крови с открытой венозной системой, чтобы избавить конечность от большого количества вектора насколько возможно. Мы полагаем, что у предварительного прилива крови могло быть особое значение в клиническом испытании. Даже в отсутствие существующего ранее иммунитета, поставка вируса в среде, свободной от серологического материала (белки комплемента, тромбоциты, эритроциты, лейкоциты), с его потенциальной вирусно-связывающей способностью, обеспечивает более легкий путь к трансдукции мышцы и снижению риска для переноса вируса к отдаленным органам. Кроме того, длительное время с отсутствием кровообращения в присутствии тромбоцитов и других факторов свертывающей системы крови могло привести к активации свертывающей системы в конечности с потенциальными серьезными последствиями.
У мышей данный подход заканчивался 80 % к 90%-ой трансдукцией мышцы, используя rAAV6 и rAAV8, несущими микродистрофин. В попытке избежать акцента на успехе исследований у мышей, мы расширяли наши исследования до исследований у приматов, которые моделируют клиническую региональную сосудистую модель поставки, близко к людям. Таким образом у обезьяны мы непосредственно проверили доставку rAAV8, eGFP через изолированную бедренную артерию, используя условия, которые будут применимы для клинического испытания (низкое давление, низкий объем, и без фармакологических средств). Мы использовали вирусные дозы, адекватные клиническим (2 ? 1012 vg/kg), и в то же самое время достигли уровней экспрессии гена в мышцах в нижних конечностях.
Наше исследование устанавливало строгие критерии для трансгенеза с определением возможностей для клинического применения, используя подход и серотип AAV, который потенциально ускорит трансдукцию мышц, и как следствие к улучшению качества жизни у детей с миодистрофией Дюшена, с возможностью применения для лечения других мышечных дистрофий. Дополнительная работа у обезьяны требуется, особенно используя векторную конструкцию, аналогичную той, которая будет использоваться практически. Исследования у примата действительно имеют много преимуществ включая 90%-ое геномное соответствие людям. Это особенно относится к исследованиям переноса генов AAV8 у обезьян с и без существующего ранее иммунитета, исследования применения определенных иммунодепрессивных режимов. Работа, представленная здесь, прокладывает путь к исследованиям экспрессии гена у примата при условиях, которые будут адекватны клинике, используя региональную сосудистую поставку Исследования у обезьяны также определяют дальнейшую изощренность в трансгенезе, используя управляемые рентгеноконтрастные катетеры с раздуваемым баллончиком на конце для точной поставки вектора в изолированные сосудистые области с возможностью последующего удаления вируса, уменьшающего нежелательное распространения к отдаленным органам.
Ссылки:
- Emery AE: Population frequencies of inherited neuromuscular diseases—a world survey.
Neuromuscul Disord 1991, 1(1):19-29. PubMed Abstract | Publisher Full Text
- Mendell JR, Moxley RT, Griggs RC, Brooke MH, Fenichel GM, Miller JP, King W, Signore L, Pandya S, Florence J, et al.: Randomized, double-blind six-month trial of prednisone in Duchenne’s muscular dystrophy.
N Engl J Med 1989, 320(24):1592-1597. PubMed Abstract
- Aartsma-Rus A, Kaman WE, Weij R, den Dunnen JT, van Ommen GJ, van Deutekom JC: Exploring the frontiers of therapeutic exon skipping for Duchenne muscular dystrophy by double targeting within one or multiple exons.
Mol Ther 2006, 14(3):401-407. PubMed Abstract | Publisher Full Text
- McClorey G, Moulton HM, Iversen PL, Fletcher S, Wilton SD: Antisense oligonucleotide-induced exon skipping restores dystrophin expression in vitro in a canine model of DMD.
Gene Ther 2006, 13(19):1373-1381. PubMed Abstract | Publisher Full Text
- Barton-Davis ER, Cordier L, Shoturma DI, Leland SE, Sweeney HL: Aminoglycoside antibiotics restore dystrophin function to skeletal muscles of mdx mice.
J Clin Invest 1999, 104(4):375-381. PubMed Abstract | Publisher Full Text | PubMed Central Full Text
- Hamed SA: Drug evaluation: PTC-124—a potential treatment of cystic fibrosis and Duchenne muscular dystrophy.
IDrugs 2006, 9(11):783-789. PubMed Abstract
- Tinsley J, Deconinck N, Fisher R, Kahn D, Phelps S, Gillis JM, Davies K: Expression of full-length utrophin prevents muscular dystrophy in mdx mice.
Nat Med 1998, 4(12):1441-1444. PubMed Abstract | Publisher Full Text
- Bogdanovich S, Krag TO, Barton ER, Morris LD, Whittemore LA, Ahima RS, Khurana TS: Functional improvement of dystrophic muscle by myostatin blockade.
Nature 2002, 420(6914):418-421. PubMed Abstract | Publisher Full Text
- Gregorevic P, Allen JM, Minami E, Blankinship MJ, Haraguchi M, Meuse L, Finn E, Adams ME, Froehner SC, Murry CE, Chamberlain JS: rAAV6-microdystrophin preserves muscle function and extends lifespan in severely dystrophic mice.
Nat Med 2006, 12(7):787-789. PubMed Abstract | Publisher Full Text
- Gregorevic P, Blankinship MJ, Allen JM, Crawford RW, Meuse L, Miller DG, Russell DW, Chamberlain JS: Systemic delivery of genes to striated muscles using adeno-associated viral vectors.
Nat Med 2004, 10(8):828-834. PubMed Abstract | Publisher Full Text | PubMed Central Full Text
- Liu M, Yue Y, Harper SQ, Grange RW, Chamberlain JS, Duan D: Adeno-associated virus-mediated microdystrophin expression protects young mdx muscle from contraction-induced injury.
Mol Ther 2005, 11(2):245-256. PubMed Abstract | Publisher Full Text
- Yoshimura M, Sakamoto M, Ikemoto M, Mochizuki Y, Yuasa K, Miyagoe-Suzuki Y, Takeda S: AAV vector-mediated microdystrophin expression in a relatively small percentage of mdx myofibers improved the mdx phenotype.
Mol Ther 2004, 10(5):821-828. PubMed Abstract | Publisher Full Text
- Sampaolesi M, Blot S, D’Antona G, Granger N, Tonlorenzi R, Innocenzi A, Mognol P, Thibaud JL, Galvez BG, Barthelemy I, Perani L, Mantero S, Guttinger M, Pansarasa O, Rinaldi C, Cusella De Angelis MG, Torrente Y, Bordignon C, Bottinelli R, Cossu G: Mesoangioblast stem cells ameliorate muscle function in dystrophic dogs.
Nature 2006, 444(7119):574-579. PubMed Abstract | Publisher Full Text
- Cerletti M, Negri T, Cozzi F, Colpo R, Andreetta F, Croci D, Davies KE, Cornelio F, Pozza O, Karpati G, Gilbert R, Mora M: Dystrophic phenotype of canine X-linked muscular dystrophy is mitigated by adenovirus-mediated utrophin gene transfer.
Gene Ther 2003, 10(9):750-757. PubMed Abstract | Publisher Full Text
- Wang Z, Zhu T, Qiao C, Zhou L, Wang B, Zhang J, Chen C, Li J, Xiao X: Adeno-associated virus serotype 8 efficiently delivers genes to muscle and heart.
Nat Biotechnol 2005, 23(3):321-328. PubMed Abstract | Publisher Full Text
- Xiao W, Chirmule N, Berta SC, McCullough B, Gao G, Wilson JM: Gene therapy vectors based on adeno-associated virus type 1.
J Virol 1999, 73(5):3994-4003. PubMed Abstract | Publisher Full Text | PubMed Central Full Text
- Schnepp BC, Clark KR, Klemanski DL, Pacak CA, Johnson PR: Genetic fate of recombinant adeno-associated virus vector genomes in muscle.
J Virol 2003, 77(6):3495-3504. PubMed Abstract | Publisher Full Text | PubMed Central Full Text
- Goyenvalle A, Vulin A, Fougerousse F, Leturcq F, Kaplan JC, Garcia L, Danos O: Rescue of dystrophic muscle through U7 snRNA-mediated exon skipping.
Science 2004, 306(5702):1796-1799. PubMed Abstract | Publisher Full Text
- Wang Z, Kuhr CS, Allen JM, Blankinship M, Gregorevic P, Chamberlain JS, Tapscott SJ, Storb R: Sustained AAV-mediated dystrophin expression in a canine model of Duchenne muscular dystrophy with a brief course of immunosuppression.
Mol Ther 2007, 15(6):1160-1166. PubMed Abstract | Publisher Full Text
- Harper SQ, Hauser MA, DelloRusso C, Duan D, Crawford RW, Phelps SF, Harper HA, Robinson AS, Engelhardt JF, Brooks SV, Chamberlain JS: Modular flexibility of dystrophin: implications for gene therapy of Duchenne muscular dystrophy.
Nat Med 2002, 8(3):253-261. PubMed Abstract | Publisher Full Text
- Rabinowitz JE, Rolling F, Li C, Conrath H, Xiao W, Xiao X, Samulski RJ: Cross-packaging of a single adeno-associated virus (AAV) type 2 vector genome into multiple AAV serotypes enables transduction with broad specificity.
J Virol 2002, 76(2):791-801. PubMed Abstract | Publisher Full Text | PubMed Central Full Text
- Clark KR, Liu X, McGrath JP, Johnson PR: Highly purified recombinant adeno-associated virus vectors are biologically active and free of detectable helper and wild-type viruses.
Hum Gene Ther 1999, 10(6):1031-1039. PubMed Abstract | Publisher Full Text
- Palmiter RD, Sandgren EP, Avarbock MR, Allen DD, Brinster RL: Heterologous introns can enhance expression of transgenes in mice.
Proc Natl Acad Sci U S A 1991, 88(2):478-482. PubMed Abstract | Publisher Full Text | PubMed Central Full Text
- Fougerousse F, Bartoli M, Poupiot J, Arandel L, Durand M, Guerchet N, Gicquel E, Danos O, Richard I: Phenotypic Correction of alpha-Sarcoglycan Deficiency by Intra-arterial Injection of a Muscle-specific Serotype 1 rAAV Vector.
Mol Ther 2007, 15(1):53-61. PubMed Abstract | Publisher Full Text
- Gonin P, Arandel L, Van Wittenberghe L, Marais T, Perez N, Danos O: Femoral intra-arterial injection: a tool to deliver and assess recombinant AAV constructs in rodents whole hind limb.
J Gene Med 2005, 7(6):782-791. PubMed Abstract | Publisher Full Text
- Wang B, Li J, Xiao X: Adeno-associated virus vector carrying human minidystrophin genes effectively ameliorates muscular dystrophy in mdx mouse model.
Proc Natl Acad Sci U S A 2000, 97(25):13714-13719. PubMed Abstract | Publisher Full Text | PubMed Central Full Text
- Hodges BL, Taylor KM, Chu Q, Scull SE, Serriello RG, Anderson SC, Wang F, Scheule RK: Local delivery of a viral vector mitigates neutralization by antiviral antibodies and results in efficient transduction of rabbit liver.
Mol Ther 2005, 12(6):1043-1051. PubMed Abstract | Publisher Full Text
- Manno CS, Pierce GF, Arruda VR, Glader B, Ragni M, Rasko JJ, Ozelo MC, Hoots K, Blatt P, Konkle B, Dake M, Kaye R, Razavi M, Zajko A, Zehnder J, Rustagi PK, Nakai H, Chew A, Leonard D, Wright JF, Lessard RR, Sommer JM, Tigges M, Sabatino D, Luk A, Jiang H, Mingozzi F, Couto L, Ertl HC, High KA, Kay MA: Successful transduction of liver in hemophilia by AAV-Factor IX and limitations imposed by the host immune response.
Nat Med 2006, 12(3):342-347. PubMed Abstract | Publisher Full Text
- Foster K, Foster H, Dickson JG: Gene therapy progress and prospects: Duchenne muscular dystrophy.
Gene Ther 2006, 13(24):1677-1685. PubMed Abstract | Publisher Full Text
- Chirmule N, Propert K, Magosin S, Qian Y, Qian R, Wilson J: Immune responses to adenovirus and adeno-associated virus in humans.
Gene Ther 1999, 6(9):1574-1583. PubMed Abstract | Publisher Full Text
Переведено проектом МойМио
Оригинальная статья : http://www.translational-medicine.com/content/5/1/45