


Введение
Мышечная дистрофия Дюшенна (МДД) является Х-хромосомным наследственным нервно-мышечным заболеванием, которое вызывается мутациями в гене дистрофина, которые вызывают дефицит белка дистрофина, ведущий к увеличению мышечной хрупкости. Мутации генерирующие короткие, и поэтому все еще частично функциональные белки, приводят к более мягким формам миопатии, иногда с очень поздним началом, известным как миодистрофия Беккера.
MDX мыши, несущее носенс- мутацию в экзоне 23, также не производят дистрофин в своих мышцах, кроме редких ревертантных дистрофина-положительных волокон в скелетных и сердечной мышце. Исключение экзона 23 из зрелой мРНК (пропуск экзонов) у MDX мыши приводит к восстановлению экспрессии дистрофина, что было достигнуто с помощью различных антисмысловых олигонуклеотидов (AOS), среди которых наиболее перспективными являются фосфоротиоат 2′-O-метил РНК (2OMePS) и фосфородиамидат морфолино олигомер (PMO). Более длительное сопротивление эндонуклеазам и более высокое сродство к мишени способны сделать PMO особенно подходящими для применения в естественных условиях.
Кроме того, PMO могут быть легко связаны с пептидами проникновения в клетку или октагуанидиновыми группами, улучшающими эффективность системной доставки в скелетные и сердечную мышцу. В настоящее время безопасность для этих последних модифицированных соединений не была продемонстрирована у людей. В настоящее время только неизменные АО используются в клинических испытаниях. Два независимых клинических испытания в Нидерландах и Великобритании уже продемонстрировали безопасность и эффективность 2OMePS и PMO АО, направленных на экзон 51 после внутримышечной инъекции у пациентов с МДД, и их системная доставка была исследована в дальнейших клинических испытаниях (исх. 27 и NTC00844597). Недавно мы показали, что выбор эффективной схемы дозирования для введения PMO является ключевым параметром в снижении количества PMO необходимого для системной доставки и, что даже более низкая дозировка, такая как 4 еженедельные инъекции 5 мг / кг, индуцирует значительное увеличение в экспрессии дистрофина.
Тем не менее, в настоящее время, нет доклинических исследований о влиянии длительного системного применения PMO у MDX мышей, и эффективность и безопасность такого длительного лечения не доказана. В настоящем исследовании мы показали, что системная доставка низких, клинически применимых, доз РМО у MDX мышей на срок до 1 года является безопасной и улучшает состояние скелетных мышц. Важно отметить, что восстанавливается экспрессия дистрофина и частично восстанавливается функциональность и сила мышц конечностей. Эти обнадеживающие результаты подтверждают целесообразность долгосрочного применения PMO у людей, как терапии для пациентов с МДД.
Результаты
Длительное повторное лечение внутривенным PMO в дозах 50 и 5 мг / кг приводит к значительному восстановлению дистрофина в скелетных мышцах мышей MDX. Шестинедельным MDX мышам вводили РМО разбавленный в стерильном физиологическом растворе через хвостовую вену. Две дозы были испытаны в этом исследовании: 5 мг / кг и 50 мг / кг, что мы и далее указывали как низкая доза (LD) и высокая доза (HD), потому что они соответственно были ниже и выше по сравнению с самой высокой дозой, используемой в настоящее время в клиническом испытании у пациентов с МДД (20 мг / кг, NTC00844597). Схема 4 еженедельных инъекций (1 цикл) и 6 недель лечения не был принят в каждой группе мышей. Цикл 4 еженедельных инъекций повторяли два или пять раз, и животные были проанализированы через 6 недель после последней инъекции (рис Дополнительное S1). После 20 недель (двух циклов обработки), дистрофин широко экспрессируется во всех мышцах, проанализированных у мышей.
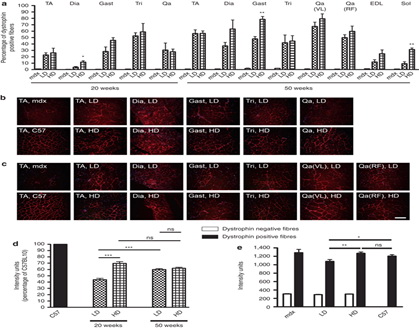
После продления лечения до 50 недель, количество дистрофин-положительных волокон оставалось неизменным в трицепсов (42 и 48% для LD и HD, соответственно, п = 4-8), тогда как оно, как правило увеличено в других протестированных мышцах по сравнению с двумя циклами. В частности, латеральная широкая и икроножная мышцы у HD-мышей содержали > 80% дистрофин-положительных волокон, тогда как более тонкие мышцы, такие как камбаловидная и длинный разгибатель пальцев, обычно экспрессируют меньший процент дистрофин-положительных волокон (рис 1а). После 20 или 50 недель лечения обе схемы доз привели к приблизительно такому же количеству дистрофин-положительных волокон в большинстве мышц, даже при том, что более сильный сигнал наблюдался после иммунофлюоресценции дистрофина в мышцах, получавших в течение 20 недель HD (рисунке 1b, в).
Полуколичественный анализ подтверждает эти наблюдения, подтверждающие, что в среднем интенсивность дистрофина в волокнах передней большеберцовой мышцы(ТА) после 20 недель лечения HD (70 ± 3% от интенсивности управления C57BL10) была выше, чем в LD мышц (44 ± 2% от контроля) (п = 4-5, p <0,0001) (рис 1г). В ТА после 50 недель участки дистрофин-отрицательных волокон были значительно снижены, но не полностью отсутствует даже при том, что общая флуоресценция выше порога, установленного для необработанных мышц, может быть результатом низкой экспрессии дистрофина в большинство волокон (рис 1в). Соответственно, после 50 недель лечения аналогичный усредненный уровень интенсивности в секциях TA, окрашенных на наличие дистрофина, была измерена для двух доз (60 ± 1,6 и 62 ± 1,5% для LD и HD по сравнению с интенсивностью C57BL10 волокна, п = 3-5, P = 0,35, рис 1г).
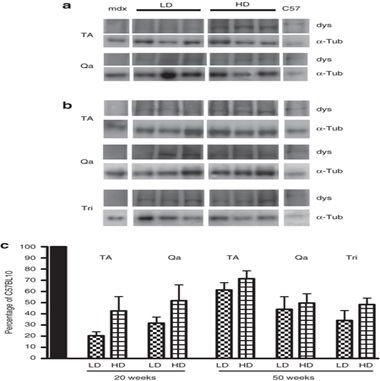
Тем не менее, когда интенсивность флуоресценции измеряли специально в дистрофин-положительных волокон этих мышц, мышечные волокна HD-мышей, породили подобное количество дистрофина по сравнению с C57BL10 волоконами (1230 ± 30 по сравнению с 1205 ± 35 единиц интенсивности, п = 5, Р = 0,16), в то время как волокна LD мышей, экспрессировали значительно меньше по сравнению с HD (1095 ± 40 единиц интенсивности) [N = 3-5, p <0,0001 (рис 1e)]. Вестерн-блот-анализ подтвердил результаты, полученные при исследовании интенсивности, показывающие, что после 20 недель лечения, мышцы TA из HD-обработанных мышей экспрессировали, как правило, больше дистрофина в мышцах LD обработанных мышей. Тем не менее, различие не было статистически значимым (20 ± 3 и 43 ± 13% от дистрофина, выраженного C57 для LD и HD, соответственно, N = 4-5, Р = 0,17).
Аналогичный результат был получен для четырехглавой мышцы (32 ± 5 и 52 ± 14% для LD и HD, соответственно, п = 4-5, P = 0,27) (рис 2а, в). После 50 недель лечения очевидное различие в уровнях экспрессии дистрофина не было найдено между LD и HD дозах в TA (61 ± 6 и 71 ± 7%, соответственно, п = 5, Р = 0,32), в четырехглавой (44 ± 11 и 50 ± 8%, N = 5, P = 0,7), и три (34 ± 9 и 48 ± 6%, N = 5, Р = 0,21) (рис 2б, в). Не наблюдалось экспрессии дистрофина в сердечной мышце мышей (данные не показаны). Эти данные свидетельствуют о том, что за 20 недель LD и HD введения РМО производится различное количество дистрофина в скелетных мышцах MDX, но это различие имеет тенденцию к снижению при более длительном введении.
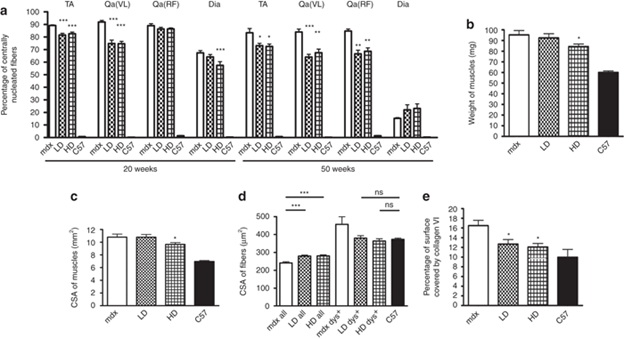
Широкая экспрессия дистрофина после длительного лечения PMO коррелирует с существенным гистологическим улучшением скелетных мышц. Значительное восстановление дистрофина наблюдается после 20 недель лечения в диафрагме HD обработанных мышей, TA и латеральной широкой мышце бедра, и соответствовала уменьшению количества волокон с центрально расположенными ядрами по сравнению с необработанными MDX мышами (в TA 82,5 ± 1,4% по сравнению с 89 ± 0,5%, в латеральной широкой 74,6 ± 1,9% по сравнению с 91,9 ± 1,3, в диафрагме после лечения HD 57,4 ± 2,9% по сравнению с 67,4 ± 1,8%, п = 4-6, P <0,0001) (рис 3а). В мышцах, представляющих меньшее количество дистрофин-положительных волокон после введения LD, таких как прямая мышца бедра (около 10-15%, не показаны на графике) и в диафрагме (3,8 ± 1,1%), не изменялось количество волокон с центральными ядрами (рис 3а).
Продолжение лечения до 50 недель снижает процент таких волокон по сравнению с мышами соответствующего возраста из группы контроля (в TA 72,4 ± 1,9 и 74 ± 1,7% для HD и LD, соответственно по сравнению с 83,3 ± 3,4% в MDX, н = 8, Р = 0,02 для LD, P = 0,012 для HD; в латеральной широкой 68 ± 1,8 и 65,3 ± 1,7% для HD и LD, соответственно по сравнению с 84,4 ± 1,2% в MDX, P = 0,0032 для LD и P <0,0001 для HD; в прямой мышцы бедра 67,5 ± 3 и 66,7 ± 2,8% для HD и LD, соответственно по сравнению с 84 ± 2,2% в MDX, P <0,0001 для LD и HD). В TA от животных, получавших HD в течение 50 недель, вес (84,2 ± 2,4 мг по сравнению с 95,3 ± 4 мг в MDX, п = 8, Р = 0,03) (рис 3b) и площадь поперечного сечения (9,6 ± 0,3 мм2 по сравнению с 10,8 ± 0,5 мм2 в MDX, п = 8, Р = 0,048) (рис 3в) значительно снизилась, что указывает на снижение типичной гипертрофии MDX. Это сопровождалось значительным увеличением средней площади поперечного сечения волокна (327 ± 10 мкм 2 и 349 ± 11 мкм 2 для LD и HD по сравнению с 270 ± 7 мкм 2 в MDX, п = 8, p <0,0001) (рис 3d).
Важно отметить, что площадь поперечного сечения дистрофин-положительных волокон достигла значений, сравнимые с обычными волокнами (Рисунок 3D), что подтверждает защитную роль экспрессии дистрофина. Наконец, фиброзная ткань в мышцах TA, определяется как коллаген VI в положительной области поперечных сечений обработанных мышц, была сравнима с таковой у нормальных мышей (12,7 ± 0,9 и 12 ± 0,7% для LD и HD по сравнению с 9,9 ± 1,6% у мышей дикого типа, п = 8, Р = 0,18 и P = 0,2 для LD и HD, соответственно), и существенно ниже, чем коллаген VI в MDX мышцах (16,5 ± 1,1%, рис 3e). Эти данные показывают, что хроническое введение низких, клинически применимых доз PMO улучшает гистологию дистрофических мышц
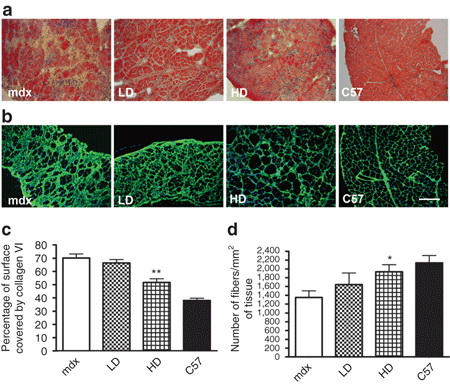
При длительном приеме PMO уменьшается фиброз и предотвращается потеря мышечных волокон в диафрагме MDX мышей.
У мышей, получавших лечение в течение 50 недель, диафрагма показала наиболее очевидное улучшение морфологической структуры особенно после обработки HD. В отличие от мышц конечностей у MDX мышей, диафрагма подвергается непрерывной дегенерации с потерей мышечных волокон начиная с 6-месячного возраста. В соответствии с этими данными, фиброзные ткани в диафрагме HD мышей занимали значительно меньшую площадь по сравнению с мышами, не получавшими лечения, как показало иммунное окрашивания на коллаген VI (51,7% от общей площади HD по сравнению с 70,2 ± 2,9% в MDX мышей, п = 8, Р = 0,0018) (рис 4 б, в).
Хотя существенной разницы не наблюдалось в волокнах с центральными ядрами между обработанными и необработанными MDX мышами (фиг.3А). Количество волокон на единицу площади диафрагмы у HD обработанных мышей была сравнима с таковой у мышей C57BL10 (1935 ± 163 волокон / мм2 в HD обработанных мышей по сравнению с 2137 ± 168 волокон / мм2 у мышей дикого типа, п = 8, Р = 0,23) (рис 4d). Эти данные показывают, что значительная потеря мышечных волокон происходит в диафрагме 13-месячных MDX мышей может быть отложена при введении клинически применимых доз РМО.
При длительном приеме PMO существенно улучшает физиологические свойства и функции MDX скелетных мышц.
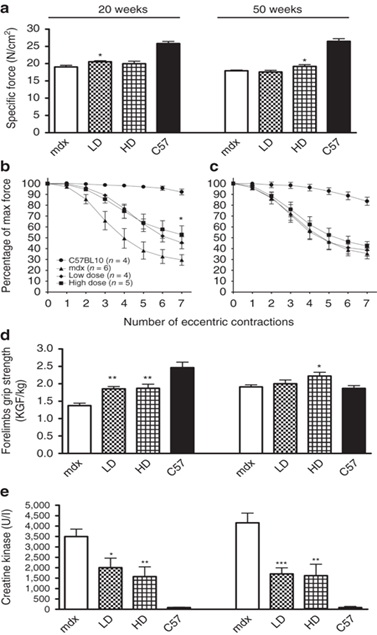
Чтобы исследовать, была ли экспрессия дистрофина в мышце LD- и HD-мышей достаточной, чтобы улучшить функции мышц, мы измерили в естественных условиях сократительные свойства мышц TA и их восприимчивость к повреждению, вызванному травмой. Мы обнаружили, что 20 недель лечения привели к восстановлению дистрофина ~ 25% в TA мышечных волокон, только привело к тенденции к улучшению силы после обработки LD [20,6 ± 0,3 Н / см2 по сравнению с 19 ± 0,4 Н / см2 в MDX, п = 4-6 (Р = 0,046)] (5а). 50 недель введения препарата вызвали значительное увеличение удельной силы ТА у HD обработанных мышей (19,2 ± 0,5 Н / см2 в HD по сравнению с 17,9 ± 0,2 Н / см2 в MDX мышей, п = 8, Р = 0,046) (рис 5а).
Интересно, что лечение MDX мышей в течение 20 недель в обеих дозах PMO индуцировало достаточные уровни дистрофина, чтобы статистически значимо улучшить устойчивость к семи эксцентричным сокращениям мышц по сравнению с MDX контроля (41,6 ± 4,9 и 50,4 ± 5,7% от первоначальной силы для LD и HD, соответственно по сравнению с 27,2 ± 3,8% в MDX мышей, п = 4-6, P <0,05) (рис 5б). Тем не менее, устойчивость к эксцентричным сокращениям не была улучшена в мышцах TA после 50 недель лечения (рис 5в).
Большое количество дистрофин-положительных волокон, наблюдаемых в трицепсе мышей, получавших в течение 20 недель лечение,коррелировало со статистически значимым увеличением силы передних конечностей обработанных мышей по сравнению с контрольными мышами MDX . Тот же тип анализа проведен после 50 недель лечения, обработанные мыши показали значительное увеличение силы конечностей у HD-обработке мышей (2,22 ± 0,11 кгс / кг для HD по сравнению с 1,91 ± 0,06 кгс / кг в MDX п = 8 , Р = 0,03) (рис 5d). Анализ концентрации креатинкиназы в крови у мышей, получавших в течение 20 недель лечение (Рисунок 5E), подтвердил улучшение функции сарколеммы (2004 ± 457 и 1570 ± 478 Ед / л для LD и HD соответственно по сравнению с 3494 ± 358 Ед / л в MDX, п = 4-6, P = 0,0322 и р = 0,0089, соответственно для LD и HD).
После пяти циклов обработки уровни креатинкиназы в сыворотке оставались на низком уровне, указывающем на функциональный эффект восстановленного дистрофина (1700 ± 192 и 1619 ± 544 Ед / л для LD и HD по сравнению с 4155 ± 468 U / л в MDX мышей, п = 8, Р = 0,0005 и р = 0,005 для LD и HD, соответственно) (рис 5e). Белковый комплекс дистрофина показал восстановление в соответствии с зависимостью доза-эффект с равномерной и широкой экспрессией в сарколемме дистрофин-положительных волокон после обработки HD (дополнительном рисунке S2b). В частности, нейрональная NO синтаза была правильно связана с сарколеммой в 80-90% дистрофин-положительных волокон (45-50% от общего количества волокон в TA, Дополнительное Рисунок S2C). Важно отметить, что никакой разницы не наблюдалось в экспрессии нейрональная NO синтазы в зависимости от количества циклов обработки. Взятые вместе, эти данные предполагают, что существует значительное функциональное улучшение в конце лечения.
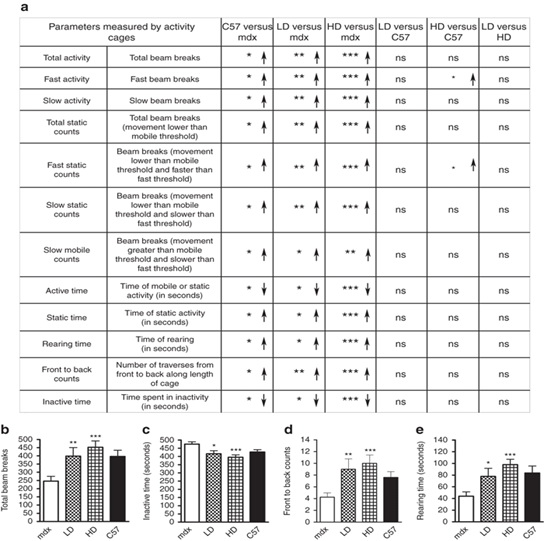
Физическая активность и состояние опорно-двигательного аппарата полностью нормализуются у MDX мышей после длительного введения PMO несмотря на только частичное восстановление уровня дистрофина.
Контроль за мышами осуществлялся с использованием теста активности открытого поля, результаты были записаны в конце эксперимента в виде 20 различных параметров (полный набор данных предоставляется по запросу). В 12 из 20 параметров 13-месячные, необработанные MDX мыши показали значительные различия в двигательном поведении по сравнению с соответствующими нормальными мышами (6а).
После пяти циклов мыши показали усиление деятельности и существенное уменьшение времени неактивности с последующей нормализацией по сравнению с соответствующими по возрасту C57BL10 мышами (Рисунок 6b-d). Большинство параметров у HD-обработанных MDX мышей имели более высокие характеристики по сравнению с C57BL10 со статистически значимыми изменениями (рис 6а). Тем не менее, 13-месячные мыши C57BL10 весили больше из-за большей жирности тела, чем mdх и это, вероятно, сократило активность мышей дикого типа. Эти данные позволяют предположить, что длительное применение РМО производит благоприятное воздействие на скелетные мышцы мышей.
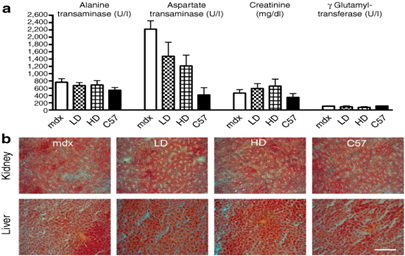
Показатели почечной и печеночной токсичности отсутствуют при длительном приеме PMO в MDX мышей В ходе эксперимента (дополнительном рисунке S3) не было обнаружено ни одного летального исхода или потери веса из-за введения PMO. Для исследования возможного токсического эффекта из-за длительного лечения концентрацию ферментов почек и печени оценивали в сыворотке. Долгосрочная доставка PMO не изменяла концентрацию аспартатаминотрансферазы, аланинаминотрансферазы, и гамма глутамилтрансферазы в сыворотке, которая предполагает, что не существует вредных эффектов РМО в печени. Концентрация креатинина была в том же диапазоне, что указывает на отсутствие негативного влияния препарата на почки (рис 7а). Гистологическое строение обоих органов также оценивали путем окрашивания гематоксилином и эозином, и не наблюдалось ни инфильтратов или вакуолизации, что подтверждало безопасность лечения (рисунок 7b). Все эти данные показали, что 50 недель лечения РМО является безопасными у MDX мышей.
Обсуждение
Выбор оптимального режима дозирования играет ключевую роль в клиническом применении любого лекарственного соединения, в том числе АО для лечения МДД. PMO является одним из наиболее перспективных химических АО из-за высокой эффективности и профиля безопасности. Однако в настоящее время, только краткосрочные исследования были проведены, и как безопасность, так и конечный результат длительного лечения до сих пор неизвестны. Мы предположили, что повторяющиеся циклы инъекций в низких дозах могут представлять эффективный режим дозирования для потенциального клинического подхода для МДД, позволяя значительное производство дистрофина во время лечения.
Как сообщалось ранее, цикл из 4 еженедельных инъекций 5 или 50 мг / кг PMO индуцировали значительные уровни дистрофина в скелетных мышцах, длящихся по крайней мере 8-10 недель. Эти PMO дозы очень малы по сравнению с крайне высокими дозами РМО, которые могут быть допущены без серьезных побочных эффектов (до 3000 мг / кг / инъекции у мышей и 320 мг / кг / в инъекции приматов). Здесь мы демонстрируем, что два цикла предлагаемого лечения индуцировали широкую и надежную экспрессию дистрофина.. Важно отметить, что мы обнаружили, что режим дозирования основывается на очень низкой, клинически применимой дозы PMO (кумулятивная доза только 40 мг / кг) значительно укрепляет мышцы, получавших MDX мышей и значительно снижает концентрацию CK в сыворотке крови.
Лечение пяти циклами дополнительно увеличило экспрессию дистрофина и улучшает дистрофический фенотип. У взрослых / пожилых MDX мышей наиболее пострадавшие мышцы диафрагмы, которые наиболее тесно повторяют суровую особенность дистрофических мышц человека. После неоднократных инъекций РМО диафрагма показала значительное улучшение гистологического строения. К началу лечения у молодых мышей РМО задержал прогрессирование патологии путем уменьшения отложения коллагена и позволяет избежать потери миофибрилл. В мышцах TA даже при том, что мы наблюдали аналогичное выражение дистрофина после обеих процедур, уровень дистрофина в волокнах HD-обработанных мышей был выше, и коррелировал с уменьшением патологии.
Эти наблюдения позволяют предположить, что более высокие дозы улучшат гистологические и функциональные результаты, обуславливающие большую защиту для волокон, чем более низкие дозы. Тем не менее, дистрофин производится в мышцах TA следующие 50 недель лечения, но это было недостаточно, чтобы защитить мышцы от повреждения после эксцентричного сжатия, хотя фиброз был сокращен после введения PMO. Наличие многих волокон с центрально расположенными ядрами показывает, что дегенерация мышц и регенерация произошла во время лечения. Вполне возможно, что прерывистый режим дозирования может привести к значительным колебаниям в экспрессии дистрофина с течением времени.
Последующая хроническая реконструкция мышцы, возможно, привела к деградации после повреждения при сокращении. Альтернативное, но не являющееся взаимоисключающее объяснение, что дозы, которые мы использовали, были достаточными для предотвращения развития патологии в относительно малоподвижных клетках MDX мышей, но не может быть достаточно, чтобы противодействовать эффектам более требовательных регулярных физических упражнений, и это может объяснить отсутствие защиты от эксцентричного сжатия.
Последнее открытие предполагает, что функциональные тесты, такие как меры защиты от эксцентричных сокращений больше, чем просто количественный метод оценки экспрессии белка, который должен быть использован для проверки лечения, основанного на восстановлении дистрофина в доклинических моделях. Это особенно актуально в случае пропуска экзона, где различные мутации связаны с производством различных дистрофинов, которые могут быть четко обнаружены в то время как потенциально связаные с очень низкой функциональностью.
Таким образом, доза PMO, возможно, будет скорректирована с учетом тяжести патологии, уровенем активности пациента и функциональностью дистрофина. Заново экспрессированый дистрофин после введения PMO был связан с глубоким совершенствованием деятельности обработанных MDX мышей. Мониторинг мышей с помощью теста открытого поля, который был недавно использован для проверки разницы в активности между мышами C57BL10 и MDX, но он никогда не был ранее использован для измерения улучшения деятельности после лечения, направленного, чтобы вызвать восстановление дистрофина.
В конце исследования мы наблюдали нормализацию параметров, связанных с горизонтальной и вертикальной активностью обработанных мышей MDX, со значениями, которые были неотличимы от C57BL10 мышей дикого типа. Особенно интригующим стало улучшение вертикальной активности, которая была недавно продемонстрирована и строго связана с устойчивостью к усталостью за счет восстановления сарколеммной локализацией нейроноеальной синтазы окиси азота у MDX мышей. Кроме того, значительное улучшение силы хвата передних конечностей указало на обобщенное значительное улучшение мышечной деятельности. Поскольку PMO не может пересечь гематоэнцефалический барьер, маловероятно, что введение PMO непосредственно влияет на поведение мышей через эффекты на уровне нервной системы. Насколько нам известно, это первый отчет, показывающий четкую нормализацию двигательной активности и двигательного поведения MDX мышей после лечения для восстановления экспрессии дистрофина.
Хотя эффективность некоторых РМО побудить пропуск экзонов в человеческой РНК или функциональность полученных форм человеческих дистрофинов, может быть относительно низкой, наши нынешние результаты показывают, что снижение экспрессии дистрофина может быть достаточным для улучшения повседневной деятельности пациентов с МДД. В соответствии с предыдущими краткосрочными исследованиями, основанными на системной доставке AO, мы не наблюдали экспрессии дистрофина в сердечных мышцах. В заключение, наши данные показывают обширное снижение дистрофической патологии у MDX мышей через 1 год при длительном приеме низких, клинически применимых доз антисмысловых РМО, предполагая, что такой подход может обеспечить безопасное и выгодное длительное лечение у пациентов с МДД.
Ссылки
England, SB, Nicholson, LV, Johnson, MA, Forrest, SM, Love, DR, Zubrzycka-Gaarn, EE et al. (1990). Very mild muscular dystrophy associated with the deletion of 46% of dystrophin. Nature 343: 180–182. | Article | PubMed | ISI | ChemPort |
Monaco, AP, Bertelson, CJ, Liechti-Gallati, S, Moser, H and Kunkel, LM (1988). An explanation for the phenotypic differences between patients bearing partial deletions of the DMD locus. Genomics 2: 90–95. | Article | PubMed | ChemPort |
Bulfield, G, Siller, WG, Wight, PA and Moore, KJ (1984). X chromosome-linked muscular dystrophy (mdx) in the mouse. Proc Natl Acad Sci USA 81: 1189–1192. | Article | PubMed | ChemPort |
Hoffman, EP, Morgan, JE, Watkins, SC and Partridge, TA (1990). Somatic reversion/suppression of the mouse mdx phenotype in vivo. J Neurol Sci 99: 9–25. | Article | PubMed | ISI | ChemPort |
Alter, J, Lou, F, Rabinowitz, A, Yin, H, Rosenfeld, J, Wilton, SD et al. (2006). Systemic delivery of morpholino oligonucleotide restores dystrophin expression bodywide and improves dystrophic pathology. Nat Med 12: 175–177. | Article | PubMed | ISI | ChemPort |
Gebski, BL, Mann, CJ, Fletcher, S and Wilton, SD (2003). Morpholino antisense oligonucleotide induced dystrophin exon 23 skipping in mdx mouse muscle. Hum Mol Genet 12: 1801–1811. | Article | PubMed | ISI | ChemPort |
Heemskerk, H, de Winter, C, van Kuik, P, Heuvelmans, N, Sabatelli, P, Rimessi, P et al. (2010). Preclinical PK and PD studies on 2′-O-methyl-phosphorothioate RNA antisense oligonucleotides in the mdx mouse model. Mol Ther 18: 1210–1217. | Article | PubMed | ISI | ChemPort |
Lu, QL, Mann, CJ, Lou, F, Bou-Gharios, G, Morris, GE, Xue, SA et al. (2003). Functional amounts of dystrophin produced by skipping the mutated exon in the mdx dystrophic mouse. Nat Med 9: 1009–1014. | Article | PubMed | ISI | ChemPort |
Lu, QL, Rabinowitz, A, Chen, YC, Yokota, T, Yin, H, Alter, J et al. (2005). Systemic delivery of antisense oligoribonucleotide restores dystrophin expression in body-wide skeletal muscles. Proc Natl Acad Sci USA 102: 198–203. | Article | PubMed | ChemPort |
Malerba, A, Thorogood, FC, Dickson, G and Graham, IR (2009). Dosing regimen has a significant impact on the efficiency of morpholino oligomer-induced exon skipping in mdx mice. Hum Gene Ther 20: 955–965. | Article | PubMed | ISI | ChemPort |
Popplewell, LJ, Trollet, C, Dickson, G and Graham, IR (2009). Design of phosphorodiamidate morpholino oligomers (PMOs) for the induction of exon skipping of the human DMD gene. Mol Ther 17: 554–561. | Article | PubMed | ISI | ChemPort |
Amantana, A and Iversen, PL (2005). Pharmacokinetics and biodistribution of phosphorodiamidate morpholino antisense oligomers. Curr Opin Pharmacol 5: 550–555. | Article | PubMed | ISI | ChemPort |
Fletcher, S, Honeyman, K, Fall, AM, Harding, PL, Johnsen, RD, Steinhaus, JP et al. (2007). Morpholino oligomer-mediated exon skipping averts the onset of dystrophic pathology in the mdx mouse. Mol Ther 15: 1587–1592. | Article | PubMed | ISI | ChemPort |
Moulton, HM, Fletcher, S, Neuman, BW, McClorey, G, Stein, DA, Abes, S et al. (2007). Cell-penetrating peptide-morpholino conjugates alter pre-mRNA splicing of DMD (Duchenne muscular dystrophy) and inhibit murine coronavirus replication in vivo. Biochem Soc Trans 35(Pt 4): 826–828. | Article | PubMed | ISI | ChemPort |
Jearawiriyapaisarn, N, Moulton, HM, Buckley, B, Roberts, J, Sazani, P, Fucharoen, S et al. (2008). Sustained dystrophin expression induced by peptide-conjugated morpholino oligomers in the muscles of mdx mice. Mol Ther 16: 1624–1629. | Article | PubMed | ISI | ChemPort |
Morcos, PA, Li, Y and Jiang, S (2008). Vivo-Morpholinos: a non-peptide transporter delivers Morpholinos into a wide array of mouse tissues. BioTechniques 45: 613–4, 616, 618 passim. | Article | PubMed | ISI
Yin, H, Moulton, HM, Seow, Y, Boyd, C, Boutilier, J, Iverson, P et al. (2008). Cell-penetrating peptide-conjugated antisense oligonucleotides restore systemic muscle and cardiac dystrophin expression and function. Hum Mol Genet 17: 3909–3918. | Article | PubMed | ISI | ChemPort |
Moulton, HM, Wu, B, Jearawiriyapaisarn, N, Sazani, P, Lu, QL and Kole, R (2009). Peptide-morpholino conjugate: a promising therapeutic for Duchenne muscular dystrophy. Ann N Y Acad Sci 1175: 55–60. | Article | PubMed | ISI
Wu, B, Li, Y, Morcos, PA, Doran, TJ, Lu, P and Lu, QL (2009). Octa-guanidine morpholino restores dystrophin expression in cardiac and skeletal muscles and ameliorates pathology in dystrophic mdx mice. Mol Ther 17: 864–871. | Article | PubMed | ISI | ChemPort |
Yin, H, Moulton, HM, Betts, C, Seow, Y, Boutilier, J, Iverson, PL et al. (2009). A fusion peptide directs enhanced systemic dystrophin exon skipping and functional restoration in dystrophin-deficient mdx mice. Hum Mol Genet 18: 4405–4414. | Article | PubMed | ISI | ChemPort |
Wu, B, Moulton, HM, Iversen, PL, Jiang, J, Li, J, Li, J et al. (2008). Effective rescue of dystrophin improves cardiac function in dystrophin-deficient mice by a modified morpholino oligomer. Proc Natl Acad Sci USA 105: 14814–14819. | Article | PubMed
Jearawiriyapaisarn, N, Moulton, HM, Sazani, P, Kole, R and Willis, MS (2010). Long-term improvement in mdx cardiomyopathy after therapy with peptide-conjugated morpholino oligomers. Cardiovasc Res 85: 444–453. | Article | PubMed | ISI
Moulton, HM and Moulton, JD (2010). Morpholinos and their peptide conjugates: Therapeutic promise and challenge for Duchenne muscular dystrophy. Biochim Biophys Acta 1798: 2296–2303. | Article | PubMed | ISI
van Deutekom, JC, Janson, AA, Ginjaar, IB, Frankhuizen, WS, Aartsma-Rus, A, Bremmer-Bout, M et al. (2007). Local dystrophin restoration with antisense oligonucleotide PRO051. N Engl J Med 357: 2677–2686. | Article | PubMed | ISI | ChemPort |
Kinali, M, Arechavala-Gomeza, V, Feng, L, Cirak, S, Hunt, D, Adkin, C et al. (2009). Local restoration of dystrophin expression with the morpholino oligomer AVI-4658 in Duchenne muscular dystrophy: a single-blind, placebo-controlled, dose-escalation, proof-of-concept study. Lancet Neurol 8: 918–928. | Article | PubMed | ISI | ChemPort |
Sazani, P, Weller, DL and Shrewsbury, SB (2010). Safety pharmacology and genotoxicity evaluation of AVI-4658. Int J Toxicol 29: 143–156. | Article | PubMed | ISI
Goemans NM, Buyse G, Tulinius M, Verschuuren JJG, de Kimpe SJ and van Deutekom JCT (2009). T.O.4 A phase I/IIa study on antisense compound PRO051 in patients with Duchenne muscular dystrophy. Neuromuscular Disorders 19: 659–660. | Article | ISI
Gregorevic, P, Blankinship, MJ, Allen, JM and Chamberlain, JS (2008). Systemic microdystrophin gene delivery improves skeletal muscle structure and function in old dystrophic mdx mice. Mol Ther 16: 657–664. | Article | PubMed | ISI | ChemPort |
Fletcher, S, Honeyman, K, Fall, AM, Harding, PL, Johnsen, RD and Wilton, SD (2006). Dystrophin expression in the mdx mouse after localised and systemic administration of a morpholino antisense oligonucleotide. J Gene Med 8: 207–216. | Article | PubMed | ISI | ChemPort |
Wu, B, Lu, P, Benrashid, E, Malik, S, Ashar, J, Doran, TJ et al. (2010). Dose-dependent restoration of dystrophin expression in cardiac muscle of dystrophic mice by systemically delivered morpholino. Gene Ther 17: 132–140. | Article | PubMed | ISI
Sazani, P, Gemignani, F, Kang, SH, Maier, MA, Manoharan, M, Persmark, M et al. (2002). Systemically delivered antisense oligomers upregulate gene expression in mouse tissues. Nat Biotechnol 20: 1228–1233. | Article | PubMed | ISI | ChemPort |
Stedman, HH, Sweeney, HL, Shrager, JB, Maguire, HC, Panettieri, RA, Petrof, B et al. (1991). The mdx mouse diaphragm reproduces the degenerative changes of Duchenne muscular dystrophy. Nature 352: 536–539. | Article | PubMed | ISI | ChemPort |
Morrison, J, Lu, QL, Pastoret, C, Partridge, T and Bou-Gharios, G (2000). T-cell-dependent fibrosis in the mdx dystrophic mouse. Lab Invest 80: 881–891. | Article | PubMed | ISI | ChemPort |
Spurney, CF, Gordish-Dressman, H, Guerron, AD, Sali, A, Pandey, GS, Rawat, R et al. (2009). Preclinical drug trials in the mdx mouse: assessment of reliable and sensitive outcome measures. Muscle Nerve 39: 591–602. | Article | PubMed | ISI
Kobayashi, YM, Rader, EP, Crawford, RW, Iyengar, NK, Thedens, DR, Faulkner, JA et al. (2008). Sarcolemma-localized nNOS is required to maintain activity after mild exercise. Nature 456: 511–515. | Article | PubMed | ISI | ChemPort |
Ishikawa, Y, Bach, JR and Minami, R (1999). Cardioprotection for Duchenne’s muscular dystrophy. Am Heart J 137: 895–902. | Article | PubMed | ISI
Markham, LW, Kinnett, K, Wong, BL, Woodrow Benson, D and Cripe, LH (2008). Corticosteroid treatment retards development of ventricular dysfunction in Duchenne muscular dystrophy. Neuromuscul Disord 18: 365–370.
Arechavala-Gomeza, V, Kinali, M, Feng, L, Brown, SC, Sewry, C, Morgan, JE et al. (2010). Immunohistological intensity measurements as a tool to assess sarcolemma-associated protein expression. Neuropathol Appl Neurobiol 36: 265–274.
Foster, H, Sharp, PS, Athanasopoulos, T, Trollet, C, Graham, IR, Foster, K et al. (2008). Codon and mRNA sequence optimization of microdystrophin transgenes improves expression and physiological outcome in dystrophic mdx mice following AAV2/8 gene transfer. Mol Ther 16: 1825–1832.
Переведено проектом МОЙМИО: www.mymio.org
Оригинал статьи: http://www.nature.com/mt/journal/v19/n2/full/mt2010261a.html
Источник МойМио


